2)在糖异生过程中,1,3-二磷酸甘油酸还原成 3-磷酸甘油醛时,需 NADH,当以乳酸为原料异生成糖时,其脱氢生成丙酮酸时已在胞液中产生了 NADH 以供利用;而以生糖氨基酸为原料进行糖异生时,NADH 则必须由线粒体内提供,可来自脂酸β-氧化或三羧酸循环。
3)甘油异生成糖耗一个 ATP,同时也生成一个 NADH 2、调节
2,6-双磷酸果糖的水平是肝内调节糖的分解或糖异生反应方向的主要信号,糖酵解加 强,则糖异生减弱;反之亦然。
3、生理意义
1)空腹或饥饿时依赖氨基酸、甘油等异生成糖,以维持血糖水平恒定。
2)补充肝糖原,摄入的相当一部分葡萄糖先分解成丙酮酸、乳酸等三碳化合物,后 者再异生成糖原。合成糖原的这条途径称三碳途径。
3)调节酸碱平衡,长期饥饿进,肾糖异生增强,有利于维持酸碱平衡。
第二章 脂类代谢
一、甘油三酯的合成代谢
合成部位:肝、脂肪组织、小肠,其中肝的合成能力最强。 合成原料:甘油、脂肪酸
1、甘油一酯途径(小肠粘膜细胞)

脂肪酸β-氧化
脂酰 CoA 进入线粒体基质后,进行脱氢、加水、再脱氢及硫解等四步连续反应,生成1 分子比原来少 2 个碳原子的脂酰 CoA、1 分子乙酰 CoA、1 分子 FADH2 和 1 分子 NADH。以上生成的比原来少 2 个碳原子的脂酰 CoA,可再进行脱氢、加水、再脱氢及硫解反应。如此反复进行,以至彻底。
4)能量生成
以软脂酸为例,共进行 7 次β-氧化,生成 7 分子 FADH2、7 分子 NADH 及 8 分子乙酰CoA,即共生成(7*2)+(7*3)+(8*12)-2=129
5)过氧化酶体脂酸氧化 主要是使不能进入线粒体的廿碳,廿二碳脂酸先氧化成较 短链脂酸,以便进入线粒体内分解氧化,对较短链脂酸无效。
三、酮体的生成和利用
组织特点:肝内生成肝外用。 合成部位:肝细胞的线粒体中。
CoA
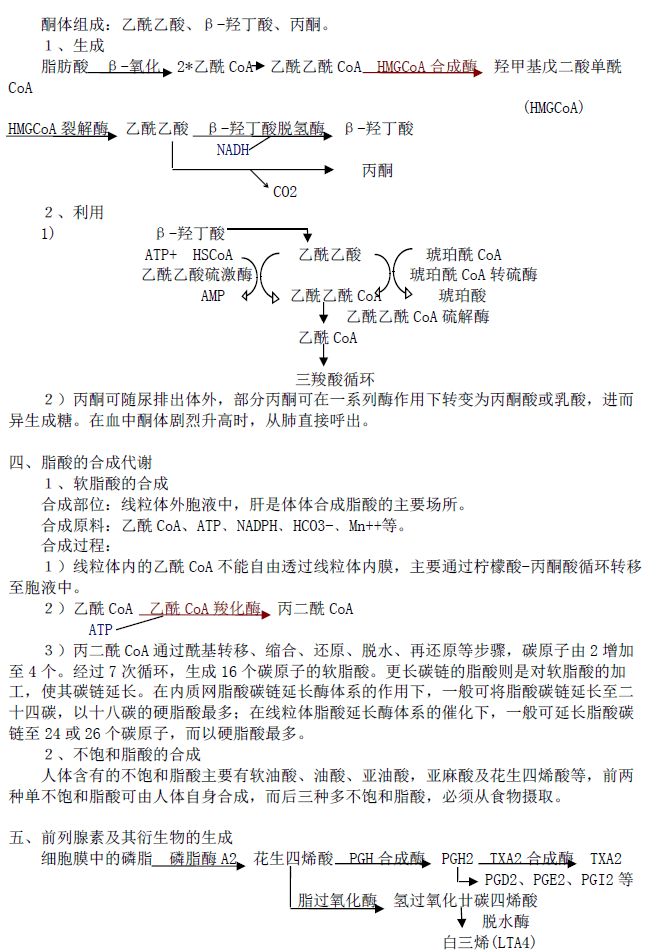
酮体组成:乙酰乙酸、β-羟丁酸、丙酮。1、生成
脂肪酸 β-氧化 2*乙酰 CoA 乙酰乙酰 CoA HMGCoA 合成酶 羟甲基戊二酸单酰
六、甘油磷脂的合成与代谢1、合成
除需 ATP 外,还需 CTP 参加。CTP 在磷脂合成中特别重要,它为合成 CDP-乙醇胺、CDP-胆碱及 CDP-甘油二酯等活化中间物所必需。
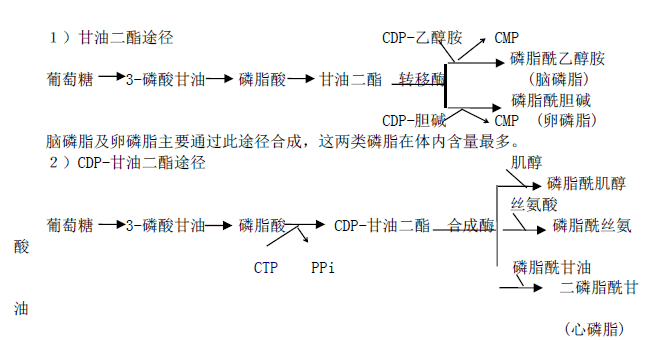
此外,磷脂酰胆碱亦可由磷脂酰乙醇胺从 S-腺苷甲硫氨酸获得甲基生成;磷脂酰丝氨
酸可由磷脂酰乙醇胺羧化生成。2、降解
生物体内存在能使甘油磷脂水解的多种磷脂酶类,根据其作用的键的特异性不同,分 为磷脂酶 A1 和 A2,磷脂酶 B,磷脂酶 C 和磷脂酶 D。
磷脂酶 A2 特异地催化磷酸甘油酯中 2 位上的酯键水解,生成多不饱和脂肪酸和溶血磷脂。后者在磷脂酶 B 作用,生成脂肪酸及甘油磷酸胆碱或甘油磷酸乙醇胺,再经甘油酸胆碱水解酶分解为甘油及磷酸胆碱。磷脂酶 A1 催化磷酸甘油酯 1 位上的酯键水解,产物是脂肪酸和溶血磷脂。
七、胆固醇代谢1、合成
合成部位:肝是主要场所,合成酶系存在于胞液及光面内质网中。
合成原料:乙酰 CoA(经柠檬酸-丙酮酸循环由线粒体转移至胞液中)、ATP、NADPH 等。合成过程:
1)甲羟戊酸的合成(胞液中)
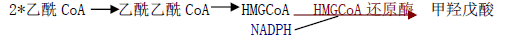
2)鲨烯的合成(胞液中)
3)胆固醇的合成(滑面内质网膜上) 合成调节:
1)饥饿与饱食 饥饿可抑制肝合成胆固醇,相反,摄取高糖、高饱和脂肪膳食后, 肝 HMGCoA 还原酶活性增加,胆固醇合成增加。
2)胆固醇 胆固醇可反馈抑制肝胆固醇的合成。主要抑制 HMGCoA 还原酶活性。3)激素 胰岛素及甲状腺素能诱导肝 HMGCoA 还原酶的合成,增加胆固醇的合成。胰
高血糖素及皮质醇则能抑制并降低 HMGCoA 还原酶的活性,因而减少胆固醇的合成;甲状腺素除能促进合成外,又促进胆固醇在肝转变为胆汁酸,且后一作用较强,因而甲亢时患者血清胆固醇含量反而下降。
2、转化
1)胆固醇在肝中转化成胆汁酸是胆固醇在体内代谢的主要去路,基本步骤为:
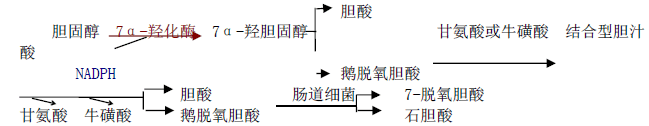
2)转化为类固醇激素 胆固醇是肾上腺皮质、睾丸,卵巢等内分泌腺合成及分泌类 固醇激素的原料,如睾丸酮、皮质醇、雄激素、雌二醇及孕酮等。
3)转化为 7-脱氢胆固醇 在皮肤,胆固醇可氧化为 7-脱氢胆固醇,后者经紫外光照射转变为维生素 D。
3、胆固醇酯的合成
细胞内游离胆固醇在脂酰胆固醇脂酰转移酶(ACAT)的催化下,生成胆固醇酯;
血浆中游离胆固醇在卵磷脂胆固醇脂酰转移酶(LCAT)的催化下,生成胆固醇酯和溶 血卵磷酯。
八、血浆脂蛋白1、分类
1)电泳法:α﹑前β﹑β及乳糜微粒
2)超速离心法:乳糜微粒(含脂最多),极低密度脂蛋白(VLDL)、低密度脂蛋白(LDL) 和高密度脂蛋白(HDL),分别相当于电泳分离的 CM﹑前β-脂蛋白﹑β-脂蛋白及α-脂蛋白等四类。
2、组成
血浆脂蛋白主要由蛋白质、甘油三酯、磷脂、胆固醇及其酯组成。乳糜微粒含甘油三 酯最多,蛋白质最少,故密度最小;VLDL 含甘油三酯亦多,但其蛋白质含量高于 CM;LDL 含胆固醇及胆固醇酯最多;含蛋白质最多,故密度最高。
血浆脂蛋白中的蛋白质部分,基本功能是运载脂类,称载脂蛋白。HDL 的载脂蛋白主要为 apoA,LDL 的载脂蛋白主要为 apoB100,VLDL 的载脂蛋白主要为 apoB﹑apoC,CM 的载脂蛋白主要为 apoC。
3、生理功用及代谢
1)CM 运输外源性甘油三酯及胆固醇的主要形式。成熟的 CM 含有 apoCⅡ,可激活脂蛋白脂肪酶(LPL),LPL 可使 CM 中的甘油三酯及磷脂逐步水解,产生甘油、脂酸及溶血磷脂等,同时其表面的载脂蛋白连同表面的磷脂及胆固醇离开 CM,逐步变小,最后转变成为 CM 残粒。
2)VLDL 运输内源性甘油三酯的主要形式。VLDL 的甘油三酯在 LPL 作用下,逐步水解,同时其表面的 apoC、磷脂及胆固醇向 HDL 转移,而 HDL 的胆固醇酯又转移到 VLDL。最后只剩下胆固醇酯,转变为 LDL。
3)LDL 转运肝合成的内源性胆固醇的主要形式。肝是降解 LDL 的主要器官。apoB100 水解为氨基酸,其中的胆固醇酯被胆固醇酯酶水解为游离胆固醇及脂酸。游离胆固醇在调节细胞胆固醇代谢上具有重要作用:①抑制内质网 HMGCoA 还原酶;②在转录水平上阴抑细胞 LDL 受体蛋白质的合成,减少对 LDL 的摄取;③激活 ACAT 的活性,使游离胆固醇酯化成胆固醇酯在胞液中储存。
4)HDL 逆向转运胆固醇。HDL 表面的 apoⅠ是 LCAT 的激活剂,LCAT 可催化 HDL 生成溶血卵磷脂及胆固醇酯。
九、高脂血症
高脂蛋白血症分型
|
分型 |
脂蛋白变化 |
血脂变化 |
|
Ⅰ |
CM↑ |
甘油三酯↑↑↑ |
|
Ⅱa |
LDL↑ |
胆固醇↑↑ |
|
Ⅱb |
LDL﹑VLDL↑ |
胆固醇↑↑甘油三酯↑↑ |
|
Ⅲ |
IDL↑ |
胆固醇↑↑甘油三酯↑↑ |
|
Ⅳ |
VLDL↑ |
甘油三酯↑↑ |
|
Ⅴ |
VLDL﹑CM↑ |
甘油三酯↑↑↑ |
注:IDL 是中间密度脂蛋白,为 VLDL 向 LDL 的过度状态。家族性高胆固醇血症的重要原因是 LDL 受体缺陷
第三章 氨基酸代谢
一、营养必需氨基酸
简记为:缬、异、亮、苏、蛋、赖、苯、色
二、体内氨的来源和转运1、来源
1)氨基酸经脱氨基作用产生的氨是体内氨的主要来源;
2)由肠道吸收的氨;即肠内氨基酸在肠道细菌作用下产生的氨和肠道尿素经细菌尿 素
酶水解产生的氨。
3)肾小管上皮细胞分泌的氨主要来自谷氨酰胺在谷氨酰胺酶的催化下水解生成的 氨。
2、转运
1)丙氨酸-葡萄糖循环
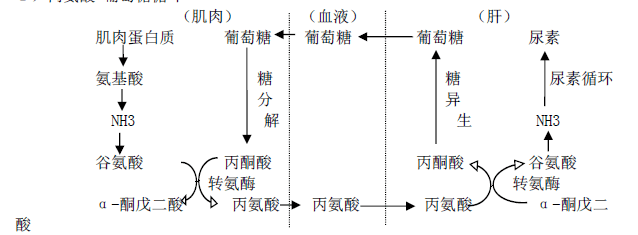
2)谷氨酰胺的运氨作用
谷氨酰胺主要从脑、肌肉等组织向肝或肾运氨。氨与谷氨酰胺在谷氨酰胺合成酶催化 下生成谷氨酰胺,由血液输送到肝或肾,经谷氨酰胺酶水解成谷氨酸和氨。
可以认为,谷氨酰胺既是氨的解毒产物,也是氨的储存及运输形式。
三、氨基酸的脱氨基作用
1、转氨基作用 转氨酶催化某一氨基酸的α-氨基转移到另一种α-酮酸的酮基上, 生成相应的氨基酸;原来的氨基酸则转变成α-酮酸。既是氨基酸的分解代谢过程,也是体 内某些氨基酸合成的重要途径。除赖氨酸、脯氨酸及羟脯氨酸外,体内大多数氨基酸可以参与转氨基作用。如:
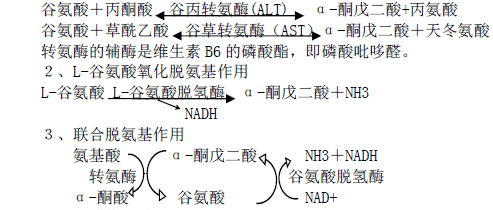
4、嘌呤核苷酸循环
上述联合脱氨基作用主要在肝、肾等组织中进行。骨骼肌和心肌中主要通过嘌呤核苷 酸循环脱去氨基。

苯酮酸尿症:当苯丙氨酸羟化酶先天性缺乏时,苯丙氨酸不能转变为酪氨酸,体内苯 丙氨酸蓄积,并经转氨基作用生成苯丙酮酸,再进一步转变成苯乙酸等衍生物。此时尿中 出现大量苯丙酮酸等代谢产物,称为苯酮酸尿症。

